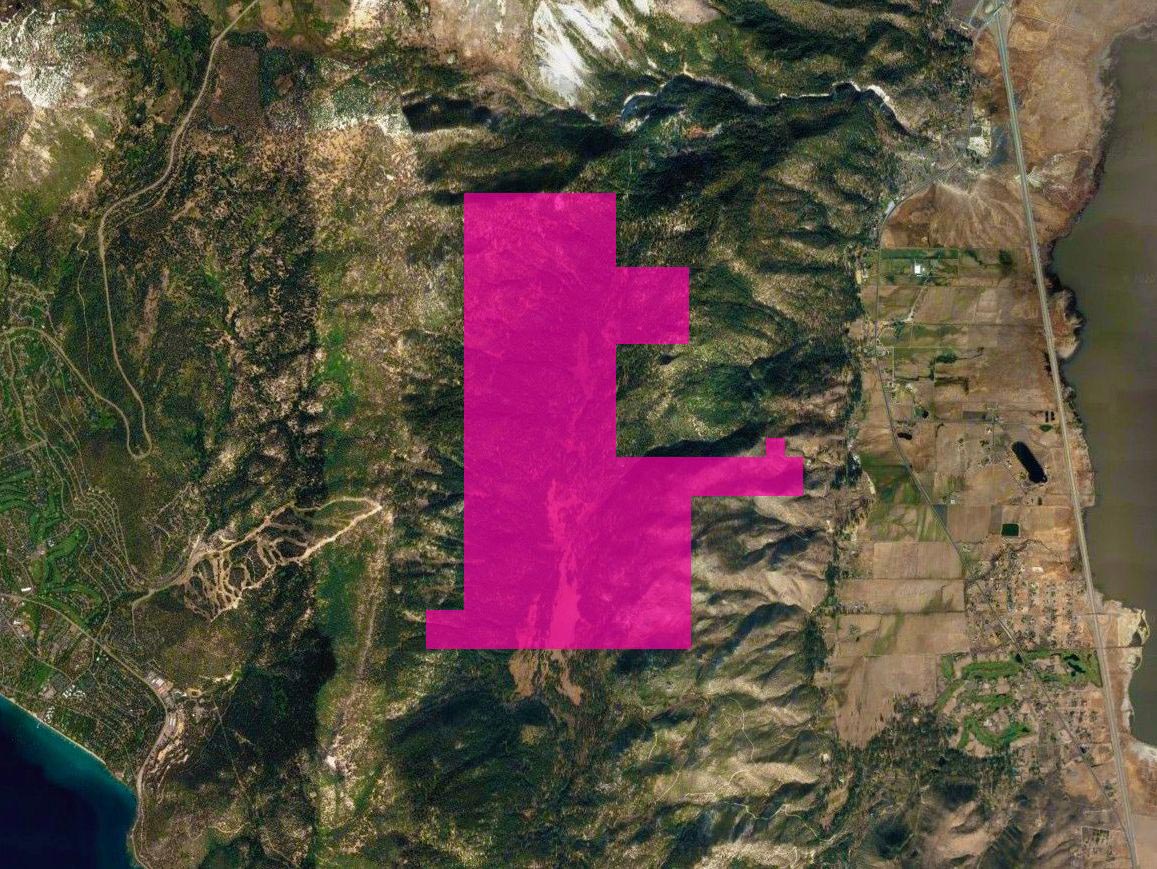
Mission
The mission of the Whittell Forest & Wildlife Area is to maintain a living field laboratory of forested mountain land for research, instruction, and community outreach that contributes to the understanding and protection of natural ecosystem integrity while engaging Nevada citizens in learning and problem-solving activities.
Land acknowledgement
We acknowledge that the University of Nevada, Reno Whittell Forest & Wildlife Area is situated on the traditional homelands of the Wašiw (Washoe) peoples. The land continues to be a gathering place for Indigenous Peoples and we recognize their deep connections to this land. We extend our appreciation for the opportunity to discover and learn on their territory.